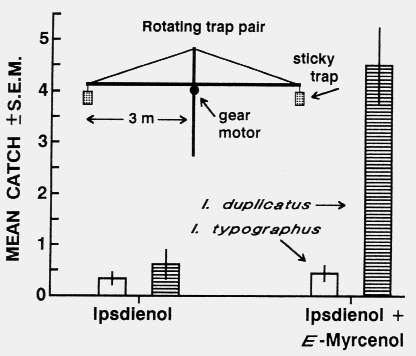
 The synergistic effect of E-myrcenol together with ipsdienol on attraction of
I. duplicatus was also demonstrated in the feld by comparison of catches on a pair of
sticky traps, one releasing the two-component blend and
the other ipsdienol alone. The two 30 cm x 30 cm diameter tubular screen traps were
coated with adhesive
(Stikem special ®) and separated 6 m apart horizontally
at a height of 1.5 m on a metal pole that was slowly
rotated by a gear motor at two revolutions per hour.
With this method each trap is exposed to all possible
paired positions during the trapping period, which minimizes the variation in catch that
would otherwise occur
with only a few flxed trap positions (traditionally the
largest component of variation [11]). Although the flying
population level of I. duplicatus was low, the attraction to
the two-component blend in the rotating trap pair was
signiflcantly higher than to ipsdienol alone (Wilcoxon
match pair test, n =18, p < 0.01; or comparison with
50:50 ratio, null hypothesis, Chi square p < 0.001,
fig. 2).
The synergistic effect of E-myrcenol together with ipsdienol on attraction of
I. duplicatus was also demonstrated in the feld by comparison of catches on a pair of
sticky traps, one releasing the two-component blend and
the other ipsdienol alone. The two 30 cm x 30 cm diameter tubular screen traps were
coated with adhesive
(Stikem special ®) and separated 6 m apart horizontally
at a height of 1.5 m on a metal pole that was slowly
rotated by a gear motor at two revolutions per hour.
With this method each trap is exposed to all possible
paired positions during the trapping period, which minimizes the variation in catch that
would otherwise occur
with only a few flxed trap positions (traditionally the
largest component of variation [11]). Although the flying
population level of I. duplicatus was low, the attraction to
the two-component blend in the rotating trap pair was
signiflcantly higher than to ipsdienol alone (Wilcoxon
match pair test, n =18, p < 0.01; or comparison with
50:50 ratio, null hypothesis, Chi square p < 0.001,
fig. 2).
1Department of Animal Ecology, Lund University, SE-223 62 Lund, Sweden
Present address:

2Chemical Ecology, Göteborg University, SE-405 30 Göteborg, Sweden
3Department of Organic Chemistry, University of Hamburg, Martin-Luther-King Platz 6, D-2000 Hamburg 13, Germany