5.2 HOST PLANT SELECTION
 Bark beetles and associated beetles feeding or living in trees must
locate a suitable host from among the relatively few scattered
widely in the forest. The host tree is restricted usually to one or
a few species and in most cases the insects seek weakened, less
resistant trees, or trees that are in the initial stages of death
and decay. Thus, it is expected that species have evolved
behavioral responses to volatile host-plant chemicals that indicate
the presence of a suitable host in which reproduction can
occur.
Bark beetles and associated beetles feeding or living in trees must
locate a suitable host from among the relatively few scattered
widely in the forest. The host tree is restricted usually to one or
a few species and in most cases the insects seek weakened, less
resistant trees, or trees that are in the initial stages of death
and decay. Thus, it is expected that species have evolved
behavioral responses to volatile host-plant chemicals that indicate
the presence of a suitable host in which reproduction can
occur.
 Bark beetles emerge from the forest litter or from the brood-
tree and search for suitable hosts in a dispersal flight. The
distances and paths of these dispersal flights are poorly known and
difficult to observe. In some experiments, beetles are painted or
marked with fluorescent powder and released to be recaptured by
pheromone-baited traps: I. typographus, the major pest of Norway
spruce, Picea abies, was found to disperse up to 8 km (Botterweg,
1982) and Ips sexdentatus up to 4 km (Jactel, 1991). Other mark-
recapture studies have found Ips and T. lineatum disperse generally
downwind (Anderbrant, 1985; Salom and McLean, 1991; Jactel, 1991)
probably due to the wind-drift component, but in light winds the
flight is nondirectional (Salom and McLean, 1989). Jactel and
Gaillard (1991) flew I. typographus on rotary flight mills
connected electronically to a computer. They found that 50% of I.
typographus can fly more than 20 km and 10% more than 45 km based
on about 50 interrupted flights. In another study, the longest
continuous fight on a flight mill was 6 h and 20 min (Forsse and
Solbreck, 1985). This suggests that a few I. typographus could fly
up to 46 km at a speed of 2 m/s without resting (Byers et al.,
1989a), and even further when blown by winds. Of course, many
individuals would be attracted to hosts or attacked trees much
nearer their dispersal origin (Lindelöw and Weslien, 1986).
Bark beetles emerge from the forest litter or from the brood-
tree and search for suitable hosts in a dispersal flight. The
distances and paths of these dispersal flights are poorly known and
difficult to observe. In some experiments, beetles are painted or
marked with fluorescent powder and released to be recaptured by
pheromone-baited traps: I. typographus, the major pest of Norway
spruce, Picea abies, was found to disperse up to 8 km (Botterweg,
1982) and Ips sexdentatus up to 4 km (Jactel, 1991). Other mark-
recapture studies have found Ips and T. lineatum disperse generally
downwind (Anderbrant, 1985; Salom and McLean, 1991; Jactel, 1991)
probably due to the wind-drift component, but in light winds the
flight is nondirectional (Salom and McLean, 1989). Jactel and
Gaillard (1991) flew I. typographus on rotary flight mills
connected electronically to a computer. They found that 50% of I.
typographus can fly more than 20 km and 10% more than 45 km based
on about 50 interrupted flights. In another study, the longest
continuous fight on a flight mill was 6 h and 20 min (Forsse and
Solbreck, 1985). This suggests that a few I. typographus could fly
up to 46 km at a speed of 2 m/s without resting (Byers et al.,
1989a), and even further when blown by winds. Of course, many
individuals would be attracted to hosts or attacked trees much
nearer their dispersal origin (Lindelöw and Weslien, 1986).
Several bark beetle species (D. pseudotsugae, S.
multistriatus, T. lineatum and I. sexdentatus) may require a period
of flight exercise before they are fully responsive to pheromone or
host attractants (Graham, 1959; Atkins, 1969; Bennett and Borden,
1971; Wollerman, 1979; Choudhury and Kennedy, 1980; Jactel, 1991).
Other species such as D. ponderosae, D. brevicomis, I.
paraconfusus, Pityogenes chalcographus and T. piniperda are
responsive to semiochemicals immediately after beginning flight
(Gray et al., 1972; Byers and Wood, 1980; Byers et al., 1985,
1990a). I. typographus emerging from the duff responded to nearby
pheromone traps without the need for an extended period of flight
exercise (Lindelöw and Weslien, 1986), although experiments in the
laboratory indicate that response increases with the length of
flight exercise (Schlyter and Löfqvist, 1986). Bark beetles
generally swallow air and inflate their ventriculus before flight,
and this may function as a barometric air pressure receptor to
indicate imminent stormy weather (Lanier and Burns, 1978).
Artificial changes in the air pressure reduced responsiveness of S.
multistriatus to the host compound ŕ-cubebene and pheromone
components (Lanier and Burns, 1978).
Finding and accepting host plants by insects has been reviewed
by Miller and Strickler (1984). They present a model (their Fig.
6.1) by Dethier (1982) where the decision by the insect whether to
accept the plant is dependent on external (olfaction, vision,
mechanoreception, and gustation) stimulatory and inhibitory inputs
balanced against internal excitatory and inhibitory inputs. A
graphical, and simplified, model of host acceptance is shown in
Fig. 6 that is directly applicable to bark beetles. In this model,
as the bark beetle flies around searching for suitable host trees
(usually trees already under attack by conspecifics) they use up
energy reserves of lipids (Atkins, 1969; Thompson and Bennett,
1971) and probably become increasingly "desperate" to accept a
host. The beetle may by chance encounter several hosts during the
dispersal flight that are more or less suitable for reproduction.
The beetle will accept the host if the combination of the host
suitability and fatigue level of the beetle is above the curve
(Fig. 6);
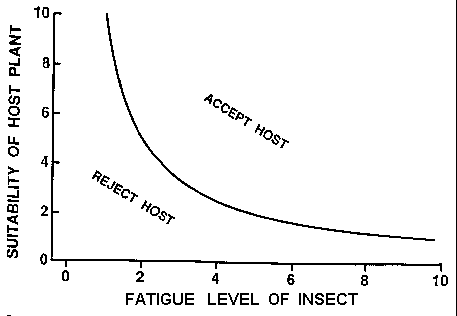
Fig. 6. Theoretical curve for the acceptance of host trees by bark
beetles depending on prerequisite flight exercise (asymptotic y-
axis), level of fatigue (amount of flight) and suitability of the
host for reproduction (which depends on nutritional quality and
density of colonization by competing bark beetles).
otherwise the beetle will continue searching for more
suitable hosts. The curve is asymptotic to the Y-axis for those
beetles that require flight before responding to semiochemicals,
whereas the curve would intersect the Y-axis for species that are
immediately responsive after emergence. The suitability of the host
is determined by the nutritional quality as well as the density of
established attacks by the same or other species of bark beetle
that indicate the potential for damaging competition.

Byers, J.A. 1995. Host tree chemistry affecting colonization in bark
beetles, in R.T. Cardé and W.J. Bell (eds.). Chemical Ecology of
Insects 2. Chapman and Hall, New York, pp. 154-213.