 Byers, J.A. 1987. Novel fraction collector for studying the oviposition rhythm in the turnip moth*.
Chronobiology International 4:189-194.
Byers, J.A. 1987. Novel fraction collector for studying the oviposition rhythm in the turnip moth*.
Chronobiology International 4:189-194.  pdf
pdf
Abstract-Turnip moths, *Agrotis segetum (Schiff.) (Lepidoptera: Noctuidae), were held in a transparent plastic box
with a window opening to a rotating paper drum upon which the females could lay eggs. A novel fraction collector,
consisting of a standard 24-hr wall timer and simple electronic circuit, served to rotate the paper in hourly
increments. The entire apparatus was housed in an environmental chamber on a 16 hr light : 8 hr dark photoperiod at
constant 23.7oC and 55% r.h. Under these conditions a circadian rhythm of egg laying was indicated with a mean
activity time 0.2 hr before dark and standard deviation of 2.1 hr. The relative egg productions during the second to
seventh day of oviposition peaked on the third day. Differences in the circadian rhythms of mating and oviposition
are considered in terms of ecological fitness.
Key words--Agrotis segetum, turnip moth, Noctuidae, circadian rhythm, oviposition, fraction collector, timer.
Introduction
In many, studies of chronobiology it is advantageous to observe the various circadian
rhythms with electronic monitors or collectors.
One useful device for observing insect rhythms
such as defecation, domicile construction, oviposition,
feeding and emergence is the multi,
test-tube holding fraction collector. While
fraction collectors are commercially available,
they are quite expensive, especially if several are
required. Byers (1) described an inexpensive
fraction collector, consisting of seven integrated
circuits, that can collect fractions of seconds,
minutes or hours with feedback positioning of
the test tubes. In many cases, however, there is no
need for such a sophisticated device that can
provide very brief or precise timing periods. A
more simple device that would produce periods
of 15, 30 or 60 min is often all that is needed to
properly evaluate many kinds of rhythms.
The electronic fraction collector described by
Byers (1) was modified in the present study by
replacing the more complicated timer circuits
with a standard 24-hr wall timer and a simple
circuit. The collector disk also was modified to
collect eggs deposited by female moths so that
oviposition rhythms could be studied under
controlled conditions.
Methods
Fraction collector
The schematic circuit of the electronic fraction
collector is shown in Figure 1.
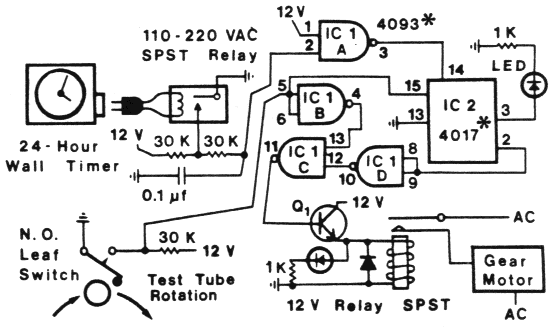
Figure 1. Schematic cirruit of the fraction collector timer (with the 24-hr wall
timer), test-tube position sensor and motor turn-on interval control. Integrated
circuits (IC) 1 and 2 have +12 V (DC) applied to pins 14 and 16, respectively, and
ground at pins 7 and 8, respectively. All resistors are 0.25 W. CMOS integrated
circuits should not be inserted into their sockets until all connections have been
made.
Although the circuit is shown to operate at 12 V (DC), any
voltage between 5 and 15 V can be used Hith an
appropriate relay,. The npn transistor, Q1 was a
2N2222 or similar and the I2-V relay, must have
contacts rated at 120-220 V. The AC operated
relay can be any type but must conform to the
house voltage since it is actuated by the 24-hr
wall timer. A normally open leaf switch with a
roller is used to sense the position of test-tubes on
the fraction collector.
The general plan of the fraction collector disk,
test-tube positioning, gear motor and leaf-switch
can be found in [Figure 4 in Ref. (1)]. The leaf
switch feedback circuit will accommodate many
different collector dimensions and gear motor
speeds. Gear motors of 2 rpm down to about 0.25
rpm will work, but slower speeds are better for
larger numbers of tubes and larger disks. For
proper operation it is crucial that the distances
between the disk axle and the ring(s) of test-tubes
be equal, otherwise the leaf switch will not
function consistently. However, spacing between
tubes around the disk periphery does not have
to be precise because of the feedback circuit.
Fraction collector operation
A 24-hr wall timer that plugs into an AC outlet
is used to actuate an AC relay that connects pin 2
of IC1A to ground. This causes the CMOS-4093
NAND Schmitt trigger to go "high" at pin 3 and
"clocks" the 5-stage Johnson decade counter
(CMOS-4017) which causes its pin 3 to go "low"
(turning off the LED) and pin 2 to go high. Both
inputs of IC1D (pins 8 and 9) thus go high
causing output pin 10 and input pin 12 to be high.
Assuming the leaf-switch is closed when in
contact with a test-tube during the non-rotating
period (1 hr in this case), then input pins 5 and 6
of IC 1B and the reset pin 15 of IC2 are connected
to ground. This allows the counter (IC2) to
advance and also causes output pin 4 of ICIB to
be high. Thus, inputs 13 and 12 of IC1C are high
when the wall timer actuates the relay and this
causes output pin 11 to go high and bias the npn
transistor into conducting current and turning
on the 12 V relay and second LED and also
switches AC power to the gear motor.
As the motor turns the fraction collector, the
test-tube moves away from the leaf-switch which
opens and allows the inputs 5 and 6 of IC1B to
go high as well as pin 15 of IC2 which resets the
counter (pin 2 goes low and pin 3 goes high).
Power to the motor continues, however, because
the inputs to IC1 C are both low causing pin 11 to
remain high. Shortly thereafter, the next test-tube
rotates into contact with the leaf-switch and
pins 5, 6 and 15 go low which causes the inputs of
IC1C both to go high and turns-off the gear
motor (pin 11 goes low). The counter is reset and
ready to advance upon the next relay pulse from
the wall timer, approximately 1 hr later.
The fraction collector with 24 test-tubes was
modified to collect moth eggs by encircling strip-chart
recorder paper around the collector disk
head such that a vertical paper tube was
constructed. The recorder paper, trimmed to
13 cm width, was taped to the two plexiglass
disks of the collector head (45 cm diam.). Thus,
discrete areas of the paper tube (about 5.8 cm
strips) could be exposed sequentially to egg-laying
females in hourly increments (controlled
by the test-tube sensing circuit and wall timer).
Oviposition rhythm
Agrotis segetum were obtained from a
laboratory culture based on insects originating
from southern Sweden and Denmark. Larvae
were reared on artificial diet (2) using potato
instead of pea beans. The culture was maintained
on a L:D 17:7 hr photoperiod at about
23-26oC.
Insects were separated by sex as pupae and
placed in containers with 1-3 adults that were fed
5% honey solution for 1-2 days. Eight females
and four males were then allowed to mate in a
5 liter bottle with paper towelling for two days
inside a walk-in environmental chamber under
L:D 16:8 hr photoperiod (about 2000 lx) with
scotophase beginning at 2200 hr and constant
23.7oC and 55% r.h. Sucrose solution (5%) in
sponge pads was available to moths during this
time and for the remainder of the experiment.
Eggs were observed on the paper towelling
beginning on the second day (July 18, 1985). The
moths then were transferred to a plexiglass
holding box in the form of an isoceles triangular
wedge that was truncated on the apex (sides 9 x
16 cm, back 9 x 21 cm, and front 9 cm high x
4 cm wide). The front apex was open and
appressed to within 1-2 mm of the rotating paper
drum/fraction collector so that females could
deposit eggs that would clear the sides of the box
during rotation but not allow moths to escape.
Eggs were scrapped off the paper and counted
each day during times when no eggs were
deposited (0700-1300).
Results and Discussion
The L:D regime was repeated for four days
(Figure 2) with maximum egg production on the
third day of oviposition.
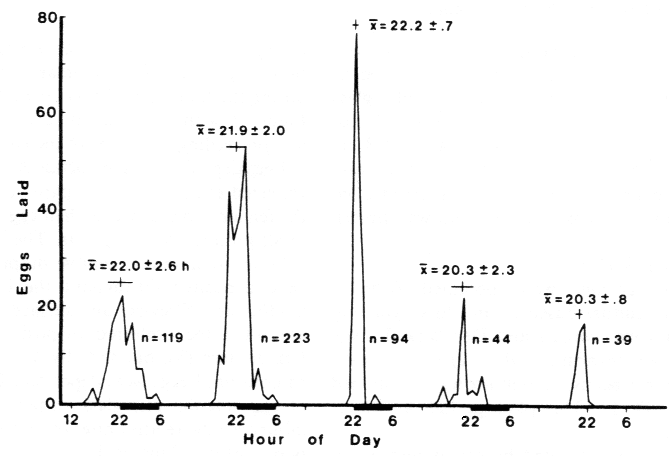
Figure 2. Eggs laid per hour on paper sections by 8 female Agrotis segetum beginning on the
third night with 4 males. Moths were held in a small plastic chamber with a 4 cm wide x 9 cm high
window appressed to a rotating paper cylinder inside an environmental chamber at constant
23.7oC and 55% r.h. and a scotophase of 8 hr as indicated. Average mean times (X bar) of egg laying
±standard deviation (±s.d.) are shown with total eggs per day given by n.
The existence of a
circadian rhythm is indicated by the peak in egg
laying on the fifth cycle under constant illumination (Figure 2).
The summary of egg laying during the four days of photoperiod is shown in
Figure 3 with the mean time of laying, 2148,
just preceeding the onset of darkness.
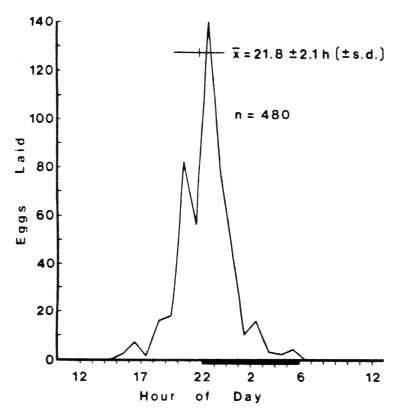
Figure 3. Total eggs laid per hour by Agrotis segetum
over a 4 day period on a L:D cycle of 16:8 hr. Average
mean time (X bar) of egg laying±standard deviation (±s.d.)
and total eggs laid (n); conditions as in Figure 2.
Eggs deposited by the noctuid A. segetum were
collected efficiently, by the paper drum because
the females normally glue their eggs, singly or in
small groups, to host plants. The egg numbers
collected represent a relative output since they
were deposited throughout the inside of the
holding box. Crevices and surface irregularities
are often the preferred ovipositional sites in
Lepidoptera (3), but initially all surfaces were
smooth until eggs were deposited. The build-up
of eggs on the plexiglass surfaces could have
influenced the relative rates of oviposition on the
paper drum. This is because the previously
deposited eggs could have acted as physical
stimuli inducing oviposition or, on the other
hand, possibly as chemical deterrents that reduce
oviposition to avoid competition. It is well
known that moths are receptive to short-range
olfactory stimuli from their host plants, and
several noctuid species are influenced by specific
host-plant compounds (4). Furthermore, there is
evidence that long-term oviposition behaviour in
some Lepidoptera is influenced by, contact with
host plants (5). In my experiments host plants
were not present so it is not known what effect
they might have on the oviposition rhythm.
The continued presence of males with the
females also could have influenced the oviposition
rhythm or egg output. In the noctuid
Earias jabia Stoll, continuous male presence
increased the egg output of the female (6). It is
also possible that males could disrupt the
oviposition behaviour with their repeated
attempts to mate. Finally, the sucrose solution
probably had an effect on, at least, egg output
since Krishna et al. (6) found that E. fabia
females produced many more eggs when fed
sugar solutions. Female noctuid moths in nature
are most certainly able to increase their egg
output by feeding on nectar from flowers.
Löfstedt et al. (7) found that virgin A. segetum
females had a "calling rhythm" of pheromone
release which had a mean activity time of
3.9 ± 0.9 hr (± s.d.) after scotophase began on a
17L:7D photoperiod. This rhythm appears to
be different from the oviposition rhythm found
in the present study which had a mean activity
time 0.2 hr before scotophase on a similar
photoperiod of 16L:8D. The zeitgebers for
these two rhythms may be different since it is
probable that the oviposition rhythm must be
timed by lights-on (16 hr earlier) while the calling
rhythm may be timed by lights-off. The zeitgeber
for the oviposition rhythm would probably not
be at lights-off as this would require a phase
angle of about 357o. The oviposition and calling
rhythms of A. segetum are similar to those of
another noctuid, Leurania separata on a 16L:8D
cycle (8). These rhythms may be independent (i.e.
different oscillators) for it is known that the
circadian rhythms of eclosion and activity in
Drosophila are functionally distinct (9).
The adaptive significance of a certain phase
angle between the LD zeitgeber and the circadian
acrophase in a particular activity can often be
generally understood. Oviposition of the diurnal
grape root borer moth, Vitacea polistiformis,
begins from 0800 to 0900 in the morning and
continues only during the daylight. Furthermore,
its mating and calling rhythms are also during
the afternoon (10). This is not surprising as the
moth uses aposematic coloration to mimic wasps
and undoubtedly derives reproductive benefits
by increasing its visual apparency. On the other
hand, noctuids are drab and camouflaged to
escape detection during the day while nocturnal
activities increase their fitness.
One of the basic tenets of biology is that a
more simple theory (one clock) is preferred over
more complex ones (two clocks) if they equally
explain the observation. Evolutionary theory
also presumes that a point can be reached where
the costs of increasing mechanistic complexity
are greater than the benefits on reproductive
fitness and thus there would be no selection
pressure to evolve further complexity. However,
there are several reasons why oviposition and
calling (mating) rhythms in A. segetum and other
noctuids may be temporally distinct. The most
obvious is that these different activities are often
mutually exclusive; but calling and then oviposition
are done at different times in the life
cycle so in theory they could have the same
rhythm. Feeding and oviposition activity would
most effectively be done at dusk or early evening
when some light remained to help in food-plant
location (both flowers and larval hosts). Also, in
terms of optimal foraging and energy consumption
it would be best to undertake feeding
and oviposition during the warmest part of the
crepuscular/nocturnal period when energy requirements
to maintain flight temperatures are
least [noctuids thermoregulate by vibrating the
thoracic wing muscles (11)]. Calling activity
could be done later in the night when light is not
essential and when colder temperatures have
little effect on energy consumption because the
females must remain sedentary and wait for
males. On the other hand, males at this time must
expend maximal energy when flying to calling
females; but then they have little choice and
besides they do not have to lay eggs [cf. (12)].
References
I. Byers J. A. Electronic fraction collector used for insect sampling in the photoperiod-induced diel emergenre of bark
beetles. Physiol. Entomol. 8, 133-138. 1983.
2. Hinks C. F. and Byers J. R Biosystematics of the genus Euxoa (Lepidoptera: Noctuidae) V. Rearing procedures and life
cycles of 36 species. (Can. Entomol. 108, 1345-1357. 1976.
3. Fenemore P. G. Oviposition of potato tuber moth, Phthorimaea operculella Zell. (Lepidoptera: Gelechiidae): the
physical nature of the oviposition. N.Z.J. Zool. 5, 591-599, 1978.
4. Burnett W. C., Jones S. B. Jr. and Mabry T. J. Influence of sesquiterpene lactones of Vernonia (Compositae) on
oviposition preferences of Lepidoptera. Am. Midl. Nat. 100, 242-246. 1978.
5. Traynier R. M. M. Long-term changes in the oviposition behaviour of the cabbage butterfly, Pieris rapae, induced by
contact with plants. Physiol. Entomol. 4, 87-96. 1979.
6. Krishna S. S., Vishwapremi K. C. and Shahi K. P. Studies on the reproduction in Earias fabia Stoll (Lepidoptera:
Nortuidae): oviposition in relation to adult nutrition, mating and some environmental factors. Entomon. 2, 11-16, 1977.
7. Löfstedt C.. Van Der Pers J. N. C., Löfqvist J., Lanne B. S., Appelgren M., Bergström G. and Thelin B. Sex Pheromone
compnnents of the turnip moth, Agrotis segetum: chemical identification, electrophysiological evaluation and
behavioural activity. J. Chem. Ecol. 10, 1305-21, 1982.
8. Kanda K. and Naito A. Behavior of oriental armyworm moth, Leucania separata, from emergence to oviposition.
Jpn. J. Appl. Entomol. Zool. 13. 69-77, 1979.
9. Engelmann W, and Mack J. Different oscillators control the circadian rhythm of eclosion and activity in Drosophila.
J. Comp. Physiol & Sens. Neural. Behav. Physiol. 127, 229-238, 1978.
10. Dutcher J. C. and All J. N. Reproductive behavior of Vitacea polistiformis Harris. J. Ga. Entomol. Soc. 13, 59-63. 1978.
11. Kammer A. E. Motor pattems during flight and warm-up in Lepidoptera. J. Exp. Biol. 48, 89-109, 1968.
12. Thornhill R. and Alcock J. The Evolution of Insect Mating Systems. Harvard University Press, Cambridge, MA. p. 547,
1983.
John A. Byers
Department of Animal Ecology, Lund University, SE-223 62 Lund, Sweden
Present address:

|
|---|