Byers, J.A. 1982. Male-specific conversion of the host plant compound, myrcene, to the pheromone (+)-ipsdienol,
in the bark beetle Dendroctonus brevicomis.
Journal of Chemical Ecology 8:363-371.  pdf
pdf
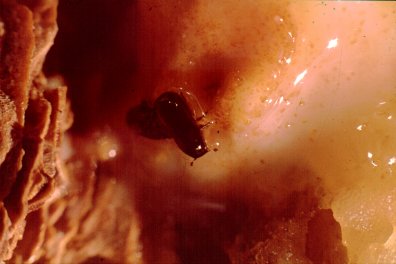 A mating pair of western pine beetles, Dendroctonus brevicomis, swimming in oleoresin of
ponderosa pine in California. The entrance hole is the darkened area just to upper right of beetle. A second beetle
is just under the most visible one.
A mating pair of western pine beetles, Dendroctonus brevicomis, swimming in oleoresin of
ponderosa pine in California. The entrance hole is the darkened area just to upper right of beetle. A second beetle
is just under the most visible one.
Abstract--
When both sexes of the bark beetle, Dendroctonus brevicomis
LeConte, were exposed to vapors of myrcene from ponderosa pine, only the
male produced (+)-ipsdienol. In the field, racemic ipsdienol significantly
reduced the attraction of both sexes in flight to a mixture of myrcene and the
aggregation pheromones, exo-brevicomin and frontalin. This suggests that
ipsdienol may be involved in regulating colonization density of D. brevicomis.
The implications of the biosynthesis of various enantiomers of
ipsdienol by D. brevicomis and the cohabitating bark beetles, Ips paraconfusus and I. pini,
in relation to their behavioral responses are discussed
in regard to reducing interspecific competition.
Key Words-
Dendroctonus brevicomis, Coleoptera, Scolytidae, Pinus
ponderosa, pheromone biosynthesis, bark beetle, myrcene, ipsdienol, exo-brevicomin, frontalin, attractants, pheromones, competition.
INTRODUCTION
The process of aggregation of the western pine beetle, D. brevicomis LeConte
(Coleoptera: Scolytidae), on a ponderosa pine, Pinus ponderosa Laws.,
begins when a female initiates the entrance hole and begins excavating a
nuptial chamber and tunnel in the phloem tissue. One component of the
aggregation pheromone, exo-brevicomin, is synthesized only in the female
during boring and feeding in the tree, is released by defecation, and causes a
low-level attraction of females and males (Silverstein et al., 1968). A male
joins the female in the nuptial chamber and immediately releases frontalin
which is synergistically attractive with exo-brevicomin (Kinzer et al., 1969;
Pitman et al., 1969; Bedard et al., 1970; Wood et al., 1976). Myrcene, a host
monoterpene, presumably is released primarily at the entrance hole from
exuding oleoresin and beetle frass, and it enhances the attraction of D. brevicomis
to the pheromones (Bedard et al., 1969, 1970).

Male (left) and female (right) D. brevicomis. Female has a "collar" or mycangium which contains
fungal spores/mycelia that are important for nutrition of larvae and for killing the pine.
Hughes (1973) reported that males of D. brevicomis produce a "large
quantity" of ipsdienol when exposed to myrcene vapors (concentration
undetermined). The production of ipsdienol is probably the result of a simple
hydroxylation of myrcene at carbon 4. However, Hughes did not mention
whether females had been exposed to myrcene. Furthermore, he indicated it
was not certain that myrcene (96%) was converted to ipsdienol since it was
"not possible to determine whether closely related monoterpenes cause the
formation of different quantities of the same products or if the small amounts
of other terpenes present as impurities are the cause." Since ipsdienol was not
known to have a behavioral effect on D. brevicomis and sex-specificity in
production was not reported, the finding until now has not been regarded as
significant.
The present study was undertaken to confirm the report of Hughes and to
determine whether females could synthesize ipsdienol. The quantities and
chirality of the ipsdienol produced in beetles after exposure to known vapor
concentrations of purified myrcene were determined. In the field, the effect of
ipsdienol on the response of D. brevicomis in flight to a mixture of its
aggregation pheromones and myrcene was tested.
METHODS AND MATERIALS
D. brevicomis adults were reared from bark removed from naturally
infested ponderosa pine trees in the Sierra National Forest, California, at
about 1000 m elevation (Browne, 1972). Both sexes of D. brevicomis were
exposed to various concentrations of myrcene vapor (GLC purified > 99.8%)
or no vapor in bottles for 18 ± 0.5 hr at 21 ± 1.7oC (Byers et al., 1979) on
several occasions under natural photoperiods (Table 1). The myrcene
concentrations were determined near the end of the exposure periods by
withdrawing headspace air from each bottle with a gas-tight syringe for
quantification by GLC analysis on a 1.8-m X 2-mm ID glass column of 3%
Apiezon L on 100/120 Gas Chrom Q at 100oC and N2 flow of 12 ml/min. The
mid- and hindguts of beetles from each bottle (Table 1) were excised and
extracted with 0.2 ml diethyl ether. The amounts of ipsdienol present in these
gut extracts were analyzed by GLC (3.6-m X 2-mm ID glass column of
Ultrabond II on 100/120 mesh at 100oC and N2 flow of 30 ml/min; and the
Apiezon L column described above). Authentic ipsdienol (GLC purified,
> 99.5%, Chem Sample Co.) was compared to gut extracts by GLC for
retention time and peak area to quantify the production in the beetles.
Eight groups of 100 males were exposed to myrcene (Table 1) as
described above and the guts of each group extracted with 0.4 ml diethyl ether
so that sufficient amounts of ipsdienol could be obtained for determination of
its optical rotation. The ipsdienol produced by these males was isolated by
injecting about 100 µl of gut extract for each GLC run and collecting the
compound from the 50:1 stream-split effluent from a 2-m X 4.5-mm ID
Teflon column of 3% Apiezon L on 100/120 Gas Chrom Q at 100oC and N2
flow of 100 ml/min. A glass U tube, 150-mm X 4-mm ID, filled 1 cm deep with
Glasperline glass beads (0.45-0.5 mm diam) and immersed in liquid N2 was
used to condense the ipsdienol from the effluent gas. After collection of
ipsdienol, the U tube was washed with ethanol and the sample rechromatographed,
collected, and washed from the U tube with a total of 300 µl of
ethanol. This sample was analyzed by GLC to determine the final concentration
of the ipsdienol collected (5.02 X 10-7 g/µl). The rotations of the
ipsdienol produced by male D. brevicomis and the authentic ipsdienol used in
the field tests were determined with an electro balancing polarimeter (Autopol
III). The ipsdienol from males and the authentic ipsdienol were further
analyzed by GCMS on a SCOT 40-m capillary column of OV-101 at 115oC.
Mass spectra (EI) were obtained with a V. G. Micromass 7070F mass
spectrometer with computerized data system.
Six trap pairs in the Sierra National Forest at about 1000 m elevation
were used to determine if ipsdienol (GLC purified > 99.5%) released from one
trap of each pair had any effect on the attraction of D. brevicomis to exo-brevicomin,
frontalin, and myrcene released at approximately equal rates
from both traps. The trap consisted of a 6-mm mesh metal-screen cylinder (19
cm diam X 30.5 cm high) coated with Stickem Special® (Bedard and Browne,
1969) placed 1.2 m above the ground. The traps of each pair were spaced 9-10
m apart and at least 100 m away from other pairs. Treatment and check were
assigned at random within each pair each day. exo-Brevicomin, frontalin, and
myrcene were each released at about 2 mg/day from glass tubes inside a glass
salt shaker (Byers and Wood, 1980). Ipsdienol was released at about 0.6
mg/day from another salt shaker containing two 4- X 65-mm glass tubes each
filled with 30 mg ipsdienol (GLC > 99.5%). The trap catches on July 1-6, 1980,
of Enoclerus lecontei (Wolcott) (Coleoptera: Cleridae), Temnochila chlorodia
(Mannerheim) (Coleoptera: Trogositidae), and both sexes of D. brevicomis
on each trap pair were compared with Wilcoxon signed rank tests (Lehmann,
1975). Sex ratio comparisons of D. brevicomis for the treatment and check
were performed by chi-square tests.
RESULTS
Female D. brevicomis did not contain detectable quantities of ipsdienol
(< 1 X 10-8 g/female) after exposure to myrcene vapors while males contained
at least 1.5 X 10-6 g ipsdienol per male after similar treatment (Table 1).
TABLE 1. Quantities of ipsdienol in male and female D. brevicomis
after an 18-hour exposure to myrcene vapor or no vapor.
|
| Date | Sex | No. exposed | Myrcene concentration
(10-6 g/ml)a | Ipsdienol per beetle
( X 10-8 g) |
|---|
| Oct. 13, 1977 | male | 20 | 4.4 ± 0.3 | 310 |
|---|
| " | male | 20 | None | 16 |
|---|
| " | female | 20 | 4.4 ± 0.3 | < 1d |
|---|
| " | female | 20 | None | < 1d |
|---|
| Nov. 16, 1977 | male | 20 | 3.9 ± 0.6 | 210 |
|---|
| " | female | 20 | 3.9 ± 0.6None | < 1d |
|---|
| May 3, 1980 | male | 30b | 2.0 ± 0.4 | 152 ± 5 |
|---|
| " | male | 30b | None | 12 ± 3 |
|---|
| " | female | 30b | 2.0 ± 0.4 | < 0.5d |
|---|
| " | female | 30b | None | < 0.5d |
|---|
| May 10, 1980 | male | 800cb | 2.1 ± 0.1 | 169 ± 9 |
|---|
aValues represent average ±SEM for all groups exposed on the same date.
bGroups of 15 per bottle.
cGroups of 100 per bottle.
dNone detected at GLC sensitivity employed.
Females unexposed to myrcene also did not contain ipsdienol; however,
unexposed males appeared to have as much as 10% of the amount of ipsdienol
that myrcene-exposed males contained (verified by GLC only). Ipsenol was
not detected in either sex. The rotation of the ipsdienol isolated by GLC from
males exposed to myrcene had a specific rotation of [alpha]D21 = +12 ± 4o
(ethanol) which is similar to [alpha]D21 = +10 ± 0.9o
reported by Silverstein et al.
(1967) for ipsdienol isolated from Ips paraconfusus Lanier (Coleoptera:
Scolytidae) [95% (+) enantiomer, Plummer et al., 1976].
The GC-MS fragmentation pattern of the purified ipsdienol obtained
from vapor-exposed males matched the mass spectra of authentic ipsdienol.
The masses of the following major fragments are listed with their percentages
of the base peak because they differ quantitatively from those published
earlier (Silverstein et al., 1967): 32:4.1, 39:3.0, 41:8.1, 53:2.8, 55:4.9, 65:2.8,
67:7.7, 68:4.9, 69:2.4, 79:5.0, 85:100.0 (base peak), 91:8.2, 109:4.6, 119:6.5,
134:5.1, 152:2.7. The quantitative differences in the spectra might be the
result of some thermal decomposition of ipsdienol on the GLC column used
by Silverstein et al.
The number of D. brevicomis caught on traps releasing aggregation
components and ipsdienol (191, 93 male : 98 female) was significantly less than
the number attracted to traps with aggregation components alone (443, 198
male : 245 female) (Figure 1).
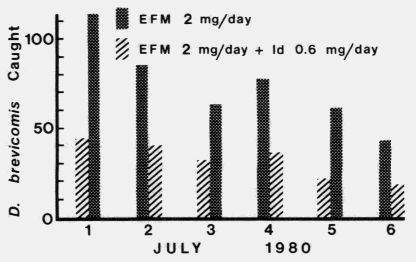
FIG. 1. The reduction in the attraction of D. brevicomis in flight to exo-brevicomin
(E), frontalin (F), and myrcene (M) by ipsdienol (Id) as shown by the comparison of
daily catch totals on 6 pairs of traps from July 1 to 6, 1980, in the Sierra National
Forest, California. The catch totals for treatment and check are shown alternating each
day to indicate random placement of ipsdienol.
The response of both sexes to the aggregation
pheromones appears to be inhibited (significantly lowered catch) by ipsdienol
(P < 0.001 in each case), and there was no significant difference between the
responses of the sexes as indicated by the ratio of catch by sex on the treatment
and check traps ( P > 0.1).

Temnochila chlorodia hunts for western pine
beetles on the bark surface and when finding one simply crushes the prey in its powerful mandiples.
The attraction of T. chlorodia, a predator, to the
aggregation components was not effected by release of ipsdienol (75 vs. 67 on
ipsdienol, P > 0.1). However, the catch of another predator, E. lecontei, was
significantly increased at traps with ipsdienol (31 vs. 5, P < 0.001) which is
consistent with their response to ipsdienol alone (Wood et al., 1966). The
rotation of the ipsdienol used in the field tests was [alpha]D21 = -0.35o (4% in
ethanol) or racemic [52% (-):48% (+)].
DISCUSSION
Myrcene appears to play a role in several important ecological functions
for D. brevicomis. The release rate of myrcene from the tree is increased by the
boring activity of beetles which use the compound in combination with
aggregation pheromones to locate its host and breeding sites. In the present
study, myrcene was converted to ( S)-(+)-ipsdienol only in the male beetle, and
racemic ipsdienol, released at rates comparable to the aggregation pheromones,
inhibited the attraction of both sexes in the field. This suggests that
ipsdienol may function in regulating the density of attack and/or terminating
the attack in an area depending on the compound's release rate in relation to
release rates of other behavioral chemicals during the colonization period.
The precise function of ipsdienol remains to be elucidated. Another role that
myrcene may play in the chemical ecology of D. brevicomis was shown by
Byers and Wood (1981a) in which at least one of the myrcene-derived
pheromones of a cohabitating bark beetle, I. paraconfusus, ipsenol and
ipsdienol (Hughes, 1974; Byers et al., 1979; Hendry et al., 1980), synergized
with another pheromonal component, cis-verbenol, to inhibit the response of
D. brevicomis to its pheromones in the laboratory. The response of D. brevicomis
to pheromone from naturally infested logs was inhibited by the
presence of logs infested with I. paraconfusus males (Byers and Wood, 1980).
These results indicated that ipsdienol/ipsenol plus cis-verbenol from I. paraconfusus
may function to reduce interspecific competition for food and
space.
In California, I. pini (Say), I. paraconfusus, and D. brevicomis may all
compete for food and space on the same host tree. I. pini produces (-)-ipsdienol
as its primary pheromone and this inhibits the response of I. paraconfusus
to its pheromones (Light and Birch, 1979). On the other hand, I. paraconfusus
produces (+)-ipsdienol (Plummer et al., 1976), and this inhibits
the response of I. pini to its pheromone (Light and Birch, 1979). D. brevicomis
probably produces (+)-ipsdienol under natural conditions because it feeds on
phloem containing myrcene (Byers, 1981). Therefore, it remains to be
established if a sufficient quantity of (+)-ipsdienol is released by D. brevicomis
to inhibit I. pini response and thus could function to reduce interspecific
competition. Release of (+)-ipsdienol from D. brevicomis would not inhibit I. paraconfusus
since this beetle uses the enantiomer as one component of its
aggregation pheromone. However, Byers and Wood (198la) showed that
verbenone from male D. brevicomis was very effective in inhibiting the
response of I. paraconfusus to its pheromones.
It appears that only specific host monoterpenes can be converted to
specific pheromones in most bark beetles studied. For example, myrcene is
not converted to cis-verbenol in I. paraconfusus (Byers et al., 1979), and
another major monoterpene found in ponderosa pine, alpha-pinene, is not
converted to ipsdienol or ipsenol in the same insect (Renwick et al., 1976;
Byers, 1981). a-Pinene is converted to the pheromones cis-verbenol in I. paraconfusus
(Brand et al., 1975; Renwick et al., 1976; Byers, 1981) and
trans-verbenol in D. ponderosae (Pitman, l971; Hughes, 1973). Exposure of I. paraconfusus
to delta-3-carene, B-pinene, and limonene did not result in
synthesis of any of the above pheromones, but other specific compounds
(unidentified) were produced (Byers, unpublished). Similar exposure of D. frontalis
to host monoterpenes resulted in the production of compounds
specific to "a particular terpene" (Hughes, 1973).
One reason that these bark beetle species, as well as D. brevicomis (and
possibly I. pini), have evolved to use myrcene and alpha-pinene as precursors for
certain of their pheromones may be due to the variation of monoterpenes in
ponderosa pine. For example, Smith ( 1964) reported that the minimum
percentages of myrcene and alpha-pinene in the oleoresin of 64 ponderosa pines in
the central Sierra Nevada of California were higher (Myrcene the highest)
than the sometimes trace amounts of delta-3-carene, B-pinene, and limonene. In a
similar study, 369 trees located at 27 sites from southern to northern
California had much more narrow ranges of percentages of myrcene and
alpha-pinene than the other three major monoterpenes (Smith et al., 1969). Thus,
there would appear to be a selective advantage for those insects that utilize
host monoterpenes which are less variable in their distribution for the
conversion to pheromones so crucial for bark beetle survival.
Byers and Wood (1981b) found that the antibiotic, streptomycin,
inhibited the conversion of myrcene to ipsdienol and ipsenol in male I. paraconfusus,
which suggests a symbiotic microorganism within the intestine
of the beetle. The biosynthetic system in D. brevicomis appears to be different
in that only ipsdienol is produced, and it is not reduced to ipsenol. Further
work is needed to determine if microorganisms play a role in D. brevicomis.
Unequivocal proof that ipsdienol is a pheromone of D. brevicomis can
only be presented when quantitative comparisons are made between the
release of the chemical from naturally infested trees and the amounts needed
to effect behavior. It appears that ipsdienol could be useful in protecting pines
from D. brevicomis attack or in mass release applications to "confuse" the
beetle. Release of ipsdienol and verbenone (Byers and Wood, 1980) would
have the advantage of inhibiting the responses of D. brevicomis, I. paraconfusus, and I. pini
to any pine substrate infested by any one of these species in
order to suppress movement of the infestation to surrounding pines or reduce
the attack rate sufficiently for the tree to overcome the beetles with resin.
Further work is needed to determine if these inhibitors of attractive response
might even repel beetles away from a place where susceptible trees are
growlng.
Acknowledgements-
I greatly appreciate the support and use of the research laboratory of
David L. Wood, University of California, Berkeley. I am grateful to Eugene Zavarin, Forest
Products Laboratory, University of California, Berkeley, for use of his polarimeter. William F.
Haddon, USDA, Western Regional Research Center, Berkeley, allowed me to assist him in
obtaining the GC-MS data. I also would like to thank William D. Bedard and Paul E. Tilden for
the use of their Forest Service research facilities at Oakhurst, California.
REFERENCES
BEDARD, W.D., and BROWNE, L.E. 1969. A delivery-trapping system for evaluating insect
chemical attractants in nature. J. Econ. Entomol. 62:1202-1203.
BEDARD, W.D., TILDEN, P.E., WOOD, D.L., SILVERSTEIN, R.M., BROWNLEE, R.G., and RODIN,
J.O. 1969. Western pine beetle: Field response to the sex pheromone and a synergistic host
terpene, myrcene. Science 164:1284-1285.
BEDARD, W.D., SILVERSTEIN, R.M., and WOOD, D.L. 1970. Bark beetle pheromones. Science
167:1638-1639.
BRAND, J.M., BRACKE, J.W., MARKOVETZ, A.J., WOOD, D.L., and BROWNE, L.E. 1972.
Production of verbenol pheromone by a bacterium isolated from bark beetles. Nature
254:136-137.
BROWNE, L.E. 1972. An emergence cage and refrigerated collector for wood-boring insects and
their associates. J. Econ. Entomol. 65:1499-1501.
BYERS, J.A. 1981. Pheromone biosynthesis in the bark beetle, Ips paraconfusus, during feeding or
exposure to vapours of host plant precursors. Insect Biochem. 11:563-569.
BYERS, J.A., and WOOD, D.L. 1980. Interspecific inhibition of the response of the bark beetles,
Dendroctonus brevicomis and Ips paraconfusus, to their pheromones in the field. J. Chem.
Ecol. 6:149-164.
BYERS, J.A., and WOOD, D.L.1981a. Interspecific effects of pheromones on the attraction of the
bark beetles, Dendroctonus brevicomis and Ips paraconfusus in the laboratory. J. Chem.
Ecol. 7:9-18.
BYERS, J.A., and WOOD, D.L.1981b. Antibiotic-induced inhibition of pheromone synthesis in a
bark beetle. Science 213:763-764.
BYERS, J.A., WOOD, D.L., BROWNE, L.E., FISH, R.H., PIATEK, B., and HENDRY, L.B. 1979.
Relationship between a host plant compound, myrcene and pheromone production in the
bark beetle Ips paraconfusus. J. Insect Physiol. 25:477-482.
HENDRY, L.B., PIATEK, B., BROWNE, L.E., WOOD, D.L., BYERS, J.A., FISH, R.H., and HICKS,
R.A.1980. In vivo conversion of a labelled host plant chemical to pheromones of the bark
beetle Ips paraconfusus. Nature 284:485.
HUGHES, P.R., 1973. Dendroctonus: Production of pheromones and related compounds in
response to host monoterpenes. Z. Angew. Entomol. 73:294-312.
HUGHES, P.R. 1974. Myrcene: A precursor of pheromones in Ips beetles. J. Insect Physiol.
20:1271-1275.
KINZER, G. W., FENTIMAN, A.F., JR., PAGE, T.F., JR., FOLTZ, R.L., VITÉ, J.P., and PITMAN, G.B.
1969. Bark beetle attractants and field bioassay of a new compound isolated from Dendroctonus.
Nature 22:475-476.
LEHMANN, E.L.1975. Nonparametrics: Statistical Methods Based on Ranks. Holden-Day, San
Francisco, 457 pp.
LIGHT, D.M., and BIRCH, M.C. 1979. Inhibition of the attractive pheromone response in Ips paraconfusus
by (R)-(-)-ipsdienol. Naturwissenschaften 66:159-160.
PITMAN, G.B. 1971. trans-Verbenol and alpha-pinene: Their utility in manipulation of the
mountain pine beetle. J. Econ. Entomol. 64:426-430.
PITMAN, G.B., VITÉ, J.P., KINZER, G.W., önd FENTIMAN, A.F., JR. 1969. Specificity of
population-aggregating pheromones in Dendroctonus. J. Insect Physiol. 15:363-366.
PLUMMER, E.L., STEWART, T.E., BYRNE, K., PEARCE, G.T., and SILVERSTEIN, R.M. 1976.
Determination of the enantiomeric composition of several insect pheromone alcohols. J.
Chem. Ecol. 2:307-331.
RENWICK, J.A.A., HUGHES, P.R., and KRULL, I.S. 1976. Selective production of cis- and trans-verbenol
from (-) and (+) alpha-pinene by a bark beetle. Science 191:199-201.
SILVERSTEIN, R.M., RODIN, J.O., and WOOD, D.L. 1967. Methodology for isolation and
identification of insect pheromones with reference to studies on California five-spined Ips. J.
Econ. Entomol. 60:944-949.
SILVERSTEIN, R.M., BROWNLEE, R.G., BELLAS, T.E., WOOD, D.L., and BROWNE, L.E. 1968.
Brevicomin: principal sex attractant in the frass of the female western pine beetle. Science
159:889-891.
SMITH, R.H. 1964. Variation in the monoterpenes of Pinus ponderosa Laws. Science 143:
1337-1338.
SMITH, R.H., PELOQUIN, R.L., JR., and PASSOF, P.A. 1969. Local and regional variation in the
monoterpenes of ponderosa pine xylem resin. USDA For. Serv. Res. Pap. PS W 56. 10 pp.
WOOD, D.L., BROWNE, L.E., SILVERSTEIN, R.M., and RODIN, J.O. 1966. Sex pheromones of bark
beetles-I. mass production, bioassay, source, and isolation of the sex pheromone of Ips confusus (LeC.).
J. lnsect Physiol. 12:523-536.
WOOD, D.L., BROWNE, L.E., EWING, B., LINDAHL, K., BEDARD, W.D., TILDEN, P.E., MORI, K.,
PITMAN, G.B., and HUGHES, P.R. 1976. Western pine beetle: specificity among enantiomers of
male and female components of an attractant pheromone. Science 192:896-898.