Byers, J.A. Chemical Ecology of Bark Beetles. Experientia 45:271-283.  pdf
pdf
CHEMICAL ECOLOGY OF BARK BEETLES
John A. Byers
Dept. of Ecology, Animal Ecology, University of
Lund, S-223 62 Lund, Sweden.
Present address: 
Summary: The purview of chemical ecology and the recent
criticisms of improper application of theory to bark beetle phenomena is briefly
discussed. Seven levels of research in chemical ecology are presented as well as
their relationship to research on bark beetles. The biology and chemical ecology of
several pest bark beetles from North America and Europe are discussed in regard
to host-tree selection theories of random landing on trees or attraction to
semiochemicals. The diversity and similarities of pheromone components among
species are presented in relation to their biosynthesis from host-tree precursors and
in relation to the ecological implications of de novo or precursor syntheses.
Individual variation in biosynthesis of, response to, and release of pheromones is
discussed. Olfactory perception of semiochemicals at both the electrophysiological
and behavioral levels is presented. Orientation to semiochemicals during walking
and flying is discussed with reference to the significance of dose-response curves
for determining a compound's functionality in short- or long-range communication.
The regulation of attack density, termination of the aggregation, mechanisms of
attack spacing, and recognition of host suitability are presented in the context of an
individual's avoidance of intra- and interspecifc competition. Finally, a brief summary
of topics where our understanding of the chemical ecology of bark beetles and their
associates is poorly known is presented.
Key words. Scolytidae, semiochemical, pheromone, allomone,
kairomone.
Chemical ecology and ecology
Ecology is the science of the relations of an organism to both its biotic and abiotic
environment which influence the organisms' distribution and
abundance77. The biotic factors are included in the disciplines
of physiology, behavior, genetics and evolution; ecology is especially concerned with
the interface of these areas. Chemical ecology then concerns any aspect of ecology
but involves the external chemicals which mediate the interactions. This definition
is actually more comprehensive than many would accept, for instance, it could
include macrophage antibody - microbe interactions, nutrient cycling, and much of
biology. Traditionally, chemical ecology has been restricted to studies of the
chemicals (semiochemicals) which mediate interactions between individuals of a
species (pheromones) or between co-evolved species (allelochemicals, such as
kairomones and allomones).
Alcock1 criticized several bark beetle (Scolytidae) researchers for what
he thought was their inadequate application of ecological theory in the explanation
of bark beetle phenomena. His primary concern was that bark beetle mass-attack
and colonization of host trees was often misunderstood in terms of "species-
selection" when in fact "individual-selection" was now the dominant theory. Today
most realize that a deeper knowledge of bark beetle biology can be obtained if both
the proximate and ultimate causes for the phenomena are considered.
Levels of research in chemical ecology
Research in chemical ecology of insects can be described with seven hierarchial
levels which to some extent are chronological. The following examples will refer to
the western pine beetle, Dendroctonus brevicomis, which aggregates en
masse in response to an aggregation pheromone. The first step in the research is
to (1) observe the biological phenomenon - e.g. demonstrate that D.
brevicomis are attracted to odor from the infested host
tree89. Once the biology is partially understood, one can design
bioassays which are used to (2) isolate and identify at least one semiochemical
component - e.g. exo-brevicomin was isolated from female frass by solvent
extraction, concentration, gas-liquid chromatographic (GC) fractionation (collecting
into discrete fractions the continuous successive elution of chemicals from the GC)
and bioassay (testing each of the several fractions for attractive
activity)129. Once a relevant compound is isolated,
enough must be obtained for structure elucidation via GC-mass spectrometry (GC-
MS), and often other spectrographic methods127-
129. The third step involves (3) isolating and identifying
all participating semiochemical components. For example, D. brevicomis
males were shown to produce frontalin73 which was synergistic
with exo-brevicomin (i.e. neither compound alone is very attractive but
together the blend is highly attractive), and the host-tree monoterpene, myrcene,
further enhanced the
attraction11,12.
Once one chemical part of the ecological system, in this case the aggregation
pheromone, is described, then (4) other pheromones, which may also act as
kairomones or allomones, can be identified with respect to the biological
phenomena. For example, intraspecific pheromone inhibitors of attraction such as
verbenone13,109 and
trans-verbenol13,32 cause
individuals to avoid colonizing in high attack density patches and thus function in
terminating the aggregation45. To further understand the
chemical ecology, which appears to grow ever more complex, one must (5) quantify
the rates of production and release of semiochemicals over the period of
colonization. Some studies have measured the release rates of some
semiochemicals from D. brevicomis over short periods27,
and others have followed the production of pheromone components in the beetle's
guts during the colonization period45.
Once the release rates of semiochemicals are known, the (6) mechanisms of
pheromone production, olfactory perception, and the sites of biosynthesis and
perception can be investigated - these areas will be discussed later. However, these
topics become increasingly more peripheral to chemical ecology as they become
more involved in physiology, behavior, genetics and evolution. Finally, one can (7)
attempt a synthesis of the knowledge concerning known semiochemicals, release
rates, interspecific interactions and biological phenomena in order to construct a
working theory on how the semiochemicals mediate the interactions between
individuals of various sexes and species.
Chemical ecology of bark beetles
Reviews of various aspects of bark beetle chemical ecology since 1980 have
concerned semiochemicals in host selection and
colonization147, host odorants and
pheromones78, orientation23,
aggregation18, aggregation
pheromones22,23, stridulation
and pheromones117,
biosynthesis59, and competition and
semiochemicals38. These reviews cover the field in more detail
than can be done here. Instead this paper will highlight the chemical ecology of a
few economically important systems: the pine beetles of the western and southern
United States and the pine and spruce beetles of Europe in regard to selected
topics.
The diversity of bark beetles and their biology can be appreciated in the
taxonomic compilation of Wood152 and in discussion
of mating systems74. However, pest bark beetles are included
in three categories: (1) "aggressive" phloem-feeding beetles which must attack en
masse living trees and kill them in order to reproduce, (2) vectors of tree diseases
such as Dutch elm disease, and (3) wood boring beetles which weaken and stain
lumber. Beetles in all three groups utilize pheromones but much of the following
discussion will best apply to those in the first group. A generalized life cycle of these
bark beetles, including many non-pest species, is depicted in Fig. 1.
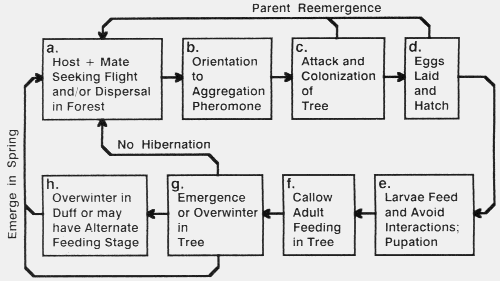
Fig. 1. Generalized life cycle of a pest bark beetle. The letters refer to the order of
discussion in the text.
Before bark beetles begin their host-tree and mate seeking flight, there may be a
period of required dispersal flight before they become responsive to
pheromone4,51,65. However, several species appear responsive to pheromone or host
attractants soon after emerging from brood
logs40,46 or
overwintering sites47.
Host selection. Little is known about dispersal and host selection except for the
stages immediately preceding landing on the tree. Host selection for several pest
species of the western United States is thought to be a random process in regard
to landing on host and non-host
trees92,153. Ponderosa
pines that were killed by freezing and screened to prohibit beetle attack, did not
exhibit higher landing rates for Ips paraconfusus, D. brevicomis or
D. ponderosae, among others, than did healthy trees. Landing rate
differences also were not observed between healthy trees and trees with diseased
roots. Other studies have shown that population densities of D.
ponderosae71 and I. typographus
3,42 are often high enough to allow nearly
every tree in a stand to be visited by at least one beetle. If beetles test the defenses
of host trees (although there is as yet no evidence of this), then, in theory, stronger
trees will repel beetles while weaker trees will allow pheromone production and
mass colonization. It is well known that conifers can be induced to exude copious
amounts of resin which can entrap
beetles45,89,8
6,125 and has some toxic
properties as well131. Healthy trees are able to exude
more resin at a higher pressure141 while weaker
trees, diseased or drought-stressed, are less able to produce resin. In late summer
when trees in California are drought-stressed, it is often observed that attacks of
D. brevicomis are not defended by resin flow and trees easily
succumb89. Thus, in many bark beetle species host selection
appears to occur after landing on the bark, and in I. paraconfusus it was
shown that a rejection of the non-host white fir did not occur until after penetration
of the phloem57.
In other species, however, there is a long-range influence of host-tree volatiles
on host selection. D. rufipennis are attracted to volatiles from white
spruce91, Scolytus multistriatus to volatiles from
American elm103, and D. valens to volatiles from
ponderosa pine139. Other bark beetles, often termed
"secondary" because they infest trees in more advanced stages of decay or are
sapwood-infesting species, have been shown to respond to volatiles from their host
trees49,64,9
0,92 and especially to
ethanol76,90. Ethanol is the most
commonly occurring kairomone attractant for the "secondary"
species76 and is supposed to be released from natural sources
due to microbial activities, although this has not been quantified. In many cases it
is known that a pheromone is released by the first arriving beetles which accounts
for most of the subsequent aggregation of the population.
Also in contrast to a random landing process during host selection, Tomicus
piniperda is strongly attracted by monoterpenes from Scots pine (terpinolene,
alpha-pinene, and 3-carene) volatilizing from wound
oleoresin47. A GC fractionation and subtractive-combination
bioassay was performed on volatile air collections of logs infested by T.
piniperda in order to find an aggregation pheromone but none was detected,
however, the host-tree monoterpenes (kairomones) were
attractive47. This phenomenon was also suggested to function
in the beetle's recognition of host susceptibility. Fallen trees or broken tops injured
during winter storms have wounds with oleoresin that release the attractive
monoterpenes and these trees are less resistant to beetles since they have a
reduced ability to produce more resin (lower turgor pressure). Ironically then, the
beetle is selecting a susceptible tree based on the very components normally used
by the tree as part of its resistance mechanism.
Once the first individuals of the "pioneer" sex arrive at the tree (males of
Ips and Pityogenes or females of Dendroctonus and
Tomicus) they are induced to begin boring into the tree. Little is known about
what chemical stimuli are involved, few feeding stimulants or deterrents have been
reported with the exception of S. multistriatus
56,63,9
7. During the development of an artificial diet for I. paraconfusus it
was found that sucrose stimulates feeding in a cellulose-based diet but not as much
as sucrose plus 22% host-phloem particles in the diet41. Other
studies have demonstrated feeding stimulant properties in extracts of host phloem
58,71. The pheromone is
released from fecal pellets after the bark beetles have fed on the phloem (and some
xylem tissue)149.
Biosynthesis and release of pheromone. Ips species
generally must feed for several hours in order to produce detectable attractants in
their fecal pellets149, and significant attraction may
not result until males have fed for more than 24
hrs149. The site of pheromone biosynthesis within
bark beetles is still uncertain but the largest amounts of semiochemicals are found
in the hindgut17,28-
32,39,69
,80,104
,107,14
2. Male Ips acquire alpha-pinene from vapor absorbed during
breathing in galleries and from ingested phloem, and then convert it to cis-
verbenol (Fig. 2) or trans-verbenol depending on whether the precursor
is of S or R
chirality75,110.
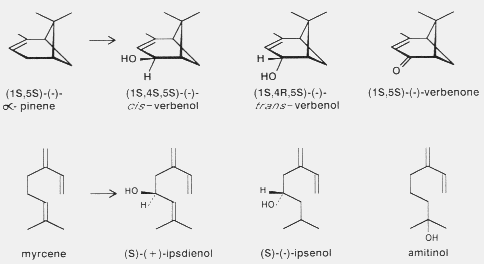
Fig. 2. Biosynthesis of the tree monoterpenes (-)-alpha-pinene to corresponding
enantiomers of cis-verbenol (an attractant pheromone component in
Ips)110,150, trans-verbenol (an inhibitory or attractant pheromone
component in Dendroctonus)32 and possibly to verbenone (an
inhibitory pheromone, allomone, and kairomone in several genera)13,39;
and conversion of the tree monoterpene myrcene to ipsdienol (pheromone
component in Ips and Dendroctonus30, ipsenol (pheromone
component of Ips)69,150 and probably (unproven) to amitinol (a
pheromone component of I. amitinus)60.
Dendroctonus species generally convert the respective S or R enantiomers
of alpha-pinene to the corresponding enantiomers of trans-
verbenol32. In contrast to trans-verbenol, the
quantities of verbenone (Fig. 2) in male D. brevicomis are not affected by
exposure to alpha-pinene vapors32.
Myrcene is converted to pheromone components hydroxylated at carbon 4
(ipsdienol and ipsenol) in various Ips
species29,69 or to amitinol in
I. amitinus (Fig. 2)60. The bio-conversion of myrcene to
ipsdienol and ipsenol in I. paraconfusus is inhibited by the antibiotic
streptomycin while other myrcene metabolites are not quantitatively
affected.41 However, no direct evidence has been found for
microbe involvement in pheromone production. The biosynthetic enzymes appear
to vary between individuals, populations, and species since the enantiomers of
ipsenol and ipsdienol vary quantitatively and qualitatively between individuals of a
population70,130 and
between populations79 of I. pini compared to other even
larger differences between species142. D.
brevicomis30 and D.
ponderosae70 also are capable of transforming myrcene to
(+)-ipsdienol which inhibits attraction to aggregation pheromone in D.
brevicomis30. In addition to the commonly used alpha-
pinene and myrcene derived pheromone components, bark beetles in the genera
Dendroctonus, Scolytus and Trypodendron utilize bicyclic ketals and
tricyclic acetals (Fig.
3)73,87,1
04,129.
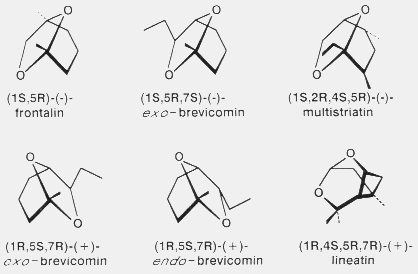
Fig. 3. Structural similarities of Dendroctonus pheromone component
enantiomers (frontalin, exo-brevicomin and endo-
brevicomin)29 to those of Scolytus multistriatus
(multistriatin)104 and Trypodendron lineatum
(lineatin)87.
In Fig. 3 it is easy to see the structural similarities of certain enantiomers of
frontalin, exo-brevicomin and multistriatin. The importance of the correct
enantiomer for each component in a pheromone was clearly demonstrated by Wood
et al.151 wherein only (-)-frontalin and (+)-exo-
brevicomin were active in causing D. brevicomis to aggregate. The
biosynthesis of these components is uncertain but appears different between the
sexes; in D. brevicomis the female produces exo-brevicomin after
feeding whereas the male contains the largest amounts of frontalin upon landing on
the tree45.
Other structures of pheromone components of bark beetles are shown in Fig.
4.
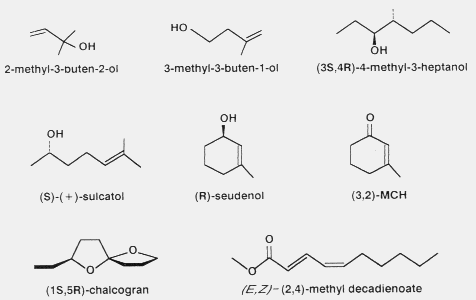
Fig. 4. Examples of other pheromone components in Ips typographus (2-
methyl-3-buten-2-ol)8, Ips cembrae (3-methyl-3-buten-1-
ol)111, Scolytus multistriatus (4-methyl-3-heptanol)104,
Gnathotrichus sulcatus (S-(+)-sulcatol)24,48, Dendroctonus
rufipennis62 and D. pseudotsugae, (R-seudenol)83,145
and (3,2-MCH)115, and Pityogenes chalcographus (S,R-
chalcogran61 and (E,Z)-(2,4)-methyl decadienoate44,46).
Short chain terpenoid compounds are found in some European species (I.
typographus, I.
cembrae)8,111 but not in the
most important American pest species. 3-Methyl-2-cyclohexen-1-one (MCH) is a
well known inhibitor of D. pseudotsugae aggregation (Fig.
4)112,115.
A spiroketal, chalcogran, is found in several species of
Pityogenes61,123, while recently a unique acetogenic pheromone component, E,Z-2,4-methyl
decadienoate (E,Z-MD), was found in P. chalcographus (Fig.
4)44,46. The biosynthesis of
beetle pheromones, with many examples for Scolytidae, has recently been reviewed
by Vanderwel and Oehlschlager138 in regard to
biosynthetic pathways (largely speculative) based on a suspected precursor. Much
of the speculation concerns the origin of the pheromone components - whether they
are terpenoid (Fig. 2), fatty acid, polyketide (Fig. 3), hydrocarbon, or amino acid (2-
phenylethanol). The physiological mechanisms of biosynthesis and hormonal control
are also reviewed. However, the ecological implications of the use of host-plant
precursors compared to the de novo synthesis of pheromone components is of
interest here.
The advantages to an individual using de novo synthesis of a pheromone
component is that control of the stereo configuration and quantity can be precisely
regulated resulting in the optimal benefit in the specific environmental context. The
possible cost is that more biosynthetic machinery is necessary with additional costs
of energy required for linking the simple building blocks. The advantages in using
an exogenous precursor in which only a small change, such as hydroxylation, is
needed to make a pheromone are obvious from an energetic standpoint. Also it
seems that alpha-pinene and myrcene are routinely de-toxified by increasing their
water solubility (hydroxylation or oxidation, Fig. 2), so the products were available
in evolutionary time to be used as pheromones. The risk for the beetle is that the
host-tree could influence the availability of monoterpene precursor or that variation
in these compounds would confer a degree of resistance to those hosts with lower
titres of precursor30. Sturgeon135
and
Smith132,133
have shown how host precursors of pheromones vary geographically in ponderosa
pine and the question is whether tree resistance might also vary due to pheromone
quantity or, as they suggest, due to toxic resistance properties of monoterpenes,
especially
limonene133,135
.
I. typographus and D. brevicomis are attracted to pheromone
blends that have varied greatly in the ratio of
components37,120. At
least in the case of I. typographus, this variation is due in part to host
precursors16,17,
75 which are not under the control of the insect.
Byers30 suggested that myrcene and alpha-pinene were utilized
by California bark beetles as precursors to pheromones because they were the
most consistently present of the monoterpenes in host trees throughout a wide
geographic range. Then, due to stabilizing-selection50
(communication requires conforming individuals) and gene flow, the species would
tend to equilibrate genetically in response to the availability of host monoterpenes,
which were de-toxified and then used as pheromone components.
The rather large variation in monoterpene precursors would, however, produce
selection pressures that would necessitate the evolution of some degree of
behavioral tolerance of variations in pheromone quantity (and component ratios) in
beetles utilizing myrcene and alpha-pinene. The inherent individual variation in
component production would be further amplified proportionally by the monoterpene
variation. Although behavioral tolerance of varying pheromone component ratios
derived exclusively from host monoterpenes have not been tested, bark beetles
seem rather more
tolerant37,120
than moths50 of pheromone component ratios.
Bark beetles even appear somewhat tolerant of variations in the ratios of
enantiomers of an attractive component. The percentages of (+)- and (-)-ipsdienol
in the natural ipsdienol of individuals in a local population of I. pini (east
Kootenay region, British Columbia) were found to vary. About half the individuals
had nearly pure (-)-enantiomer, 20% had about 10% (+), while 5% had about 35%
(+)130. Over the geographic range of I. pini,
the western population (British Columbia) averaged nearly 100% (-)-ipsdienol while
the eastern population (New York) produced about 65% (+)-
ipsdienol79. The response of the western population is inhibited
by the (+)-enantiomer, not because of an avoidance of eastern individuals but
because of avoidance of its competitor I. paraconfusus which produces (+)-
ipsdienol20. The eastern population would not be attracted to
western individuals because insufficient (+)-enantiomer is
produced20.
Few studies of bark beetles have measured the release of semiochemicals from
natural sources due to the small amounts of semiochemical relative to
contaminating host compounds. Browne et al.27 quantified the
release of attractive and inhibitory pheromone components of D. brevicomis
boring in ponderosa pine by liquefaction of air. Since then porapak Q has been
used as an adsorbent for attractive volatiles from T.
piniperda43,47,
80, I.
typographus15,122
and P. chalcographus46. This type of data is necessary
to formulate theoretical models of semiochemical interactions during colonization of
the host tree.
Observations of the immediate attraction of D. frontalis to incipiently colonized
trees under high beetle densities caused Vité et
al.142 to propose a "contact" pheromone whereby
beetles would release pheromone upon landing on an attractive tree. While there
have been no studies to determine the actual time of release, from an individual
beetle's point of view it would seem better to release pheromone after acquiring an
entrance tunnel with a female before committing resources (pheromone) to help in
the colonization of the tree. Otherwise, a beetle might lose the advantage of arriving
earlier and be outcompeted for access to females. In D. brevicomis males,
which also contain frontalin and some inhibitors in the largest amounts upon landing
(trans-verbenol, verbenone, and ipsdienol)45, it would
only make sense to an individual to release inhibitors from a place that is to be
"defended against" competitors rather than just anywhere on the tree. This also
applies for release of 3-methyl-2-buten-2-ol (MB) by male I. typographus who
contain relatively large amounts upon landing17.
Olfactory perception of semiochemicals. The chirality of semiochemicals in
bark beetles is important both in communication and in insect-plant interactions. The
enantiomeric specificity is obtained by (1) the availability of chiral precursors of the
host tree, (2) the chiral specificity of synthesizing enzymes, and (3) the enantiomer-
specific acceptors on olfactory cells55.
The electrophysiological response of an insect to chemicals can be measured
using the electroantennogram (EAG) of the whole antenna or the single-cell
technique which measures electrical responses of specific receptor cells. The
antennal receptor cells each contain multiple acceptor sites that physically interact
with the chemicals. Bark beetle olfactory cells on the antennae have been shown
to be of several functional types, all of which generally are found in each species:
(1) a "labelled-line" or highly specific receptor cell such as the ipsdienol-sensitive
cells in I. paraconfusus and I. pini which are responsive only to one
or the other enantiomer94, (2) a pheromonal cell which is also
responsive to some other synergists or inhibitors such as the frontalin cells of D.
frontalis (the cells have at least two acceptor types specific for each enantiomer of
frontalin)54,102, and
(3) "generalist" receptor cells which respond to host monoterpenes as well as
pheromones to some extent54,55.
However, the possibility that impurities in the monoterpenes account for the receptor
activity has not been definitively ruled out.
The technique of "differential adaptation", in which a receptor cell (or antenna) is
first adapted with a compound by saturation of specific acceptor sites and then
exposed to a different volatile to see if an electrical response can be elicited, has
been used to determine the types of semiochemical acceptors on a
cell55,99,1
00. By using this technique with single-cell recordings it has
been shown that D. pseudotsugae has at least four olfactory cell
types55. Three types are each most sensitive to either MCH,
seudenol, or frontalin, although they are all stimulated somewhat by all of these
pheromone components55. The fourth cell type is most
sensitive to beetle-produced synergists and host attractants and is less sensitive to
pheromone components55. Acceptors are specific for one
enantiomer of a chiral mixture101 and there may be
either only one type of acceptor per cell (e.g. (+)- or (-)-ipsdienol in I.
paraconfusus94) or both types of chiral acceptors on the
same cell (e.g. (+)- and (-)-frontalin in D. frontalis102
and D. pseudotsugae55).
In I. paraconfusus, both sexes have equal receptor sensitivity (EAG) to
natural pheromone and to (+)-ipsdienol85. However, the males
have been shown to be relatively less attracted by higher concentrations of synthetic
pheromone components and were also not as likely to fly directly to the pheromone
source as were females33. Thus, the sexual differences in
behavioral response appear to be the result of differences in central nervous system
(CNS) integration. Both sexes of I. typographus have similar dose-response
curves for cis-verbenol but some differences for MB53.
Similar to I. paraconfusus, males of I. typographus also are less
directed to pheromone at the final landing than are
females121; this behavioral difference could be due
either to CNS differences between the sexes or to the lesser receptor sensitivity of
males to MB.
Mustaparta et al.94 have shown that eastern U.S. populations
of I. pini have separate cells for the two ipsdienol enantiomers that are
synergistic attractants79. However, the exposure of the two cells
to the enantiomers together did not synergistically increase the nerve impulse rate
so it was concluded that synergism here acts at the CNS
level94. In contrast, Dickens et al.55 have
shown that the behavioral effects of MCH on D. pseudotsugae may be due in large
part to peripheral receptors rather than as a result of CNS discrimination. At low
concentrations MCH has acted as an attractant
synergist112 and certain olfactory cells have been
found which are highly specific and sensitive to MCH in both
sexes55. The dose-response curve was judged to be relatively
wide, suggesting a long-range orientation effect. However, at higher concentrations,
MCH caused a decrease in spontaneous activity of other receptor cell types
sensitive to other components55. Thus, the behavioral inhibition
of attraction by
MCH112,115
could be the result of CNS reception of the impulse frequency of the specific
receptor type and also the peripheral inhibition of receptor
activity55. Whether this peripheral inhibition is a general
phenomenon of semiochemical inhibitors is not known, although the ipsdienol
inhibitors of Ips, mentioned above, appear to function only at the CNS level.
Of the three or four receptor types found in most
species54,55,9
4,95 it appears that individual
receptors within a type also can vary in their response spectrum to various
chemicals94,95,1
37. In the few cases so far known, the host-responsive
cells are present in both sexes but the host-selecting sex (males of Ips or females
of Dendroctonus) is slightly more sensitive to plant
compounds53-55. However, the role of plant
compounds in long-range orientation of Ips is not certain; while short-range behavior
is probably influenced, it also is poorly understood. I. typographus is
purported to be sensitive to exo-brevicomin since it increased response to
the pheromone components137, although it is not
present in this species15-17. However,
sympatric D. micans and/or Dryocoetus sp. produce exo-
brevicomin and thus I. typographus could locate susceptible host trees by
responding to species that colonize weakened
hosts137. This same phenomenon, on the behavioral
level, was suggested for D. brevicomis and I.
paraconfusus40. However, the effects of "unnatural"
semiochemicals on bark beetle receptors must be considered with caution. Lanne
et al.80 have shown that T. piniperda shows EAG
responses to exo-brevicomin, ipsenol, and other compounds not known to
be associated with the beetle or any competing species.
The difficulties with electrophysiological methods for unraveling ecological
phenomena are two-fold: (1) the electrical responses may not be correlated to the
relevant behavior and (2) the nerve impulse patterns vary even within a cell
type94, all of which are not located and tested, and the patterns
are further integrated in the "black-box" CNS. It remains a great challenge to
understand even incompletely the "cross-talk" interactions and "decoding" of nerve
impulses from receptor cells at the peripheral level, a nearly impossible challenge
when the CNS is considered.
Orientation to semiochemicals. Compared to
moths5,72, much less is known about
orientation mechanisms of flying bark beetles in part due to their small size and
difficulty of study in wind-tunnels51. A rotating wind-vane trap
has been used to confirm the belief that bark beetles fly up-wind in response to a
pheromone36, however, little more is known of flight behavior.
Byers33 used grids of traps distant from a natural pheromone
source and found that male I. paraconfusus do not fly directly to the source
as females do, the reason being that males avoid patches densely colonized with
potentially intense competition. This phenomenon has been observed in other
polygynous species, e.g. I. typographus120
and P. chalcographus44. Sex-specific differences in
D. brevicomis induced by trans-verbenol were observed when females
were found to be inhibited from entering holes in artificial hosts releasing synthetic
pheromone attractants while males were not. Sexual differences were not found in
long-range orientation to pheromone component mixtures32, but
single components (either exo-brevicomin or frontalin) caused the sexes to
respond differently35,37. The
orientation of flying bark beetles is expected to use optomotor (visual) self-steered
counterturning and anemotaxis in accordance with moth
studies5,72, although we have little
evidence to support this belief.
Orientation by walking has been studied more
often19,20,3
0,32,33
,40,46,4
7 but with the objectives of observing attraction or inhibition by
semiochemical blends. Recent studies have shown that bark beetles can orient in
still air up a diffusion gradient148. Although still air
does not normally occur in nature this chemotactic mechanism may operate a few
millimeters from an entrance hole. Host monoterpenes (camphene, alpha-pinene,
á-pinene) of Norway spruce caused P. chalcographus to enter "beetle-sized"
holes in cylindrical "tree-sized" traps more so than controls (both released
pheromone components also)44.
It was proposed that a long-range orientation function for a semiochemical was
indicated if the compound had both a low threshold and a wide range of
concentrations which elicited an electrophysiological response using EAG or single-
cell recording. Conversely, a short-range function was indicated for a semiochemical
if the electrophysiological activities were observed only with a relatively high
threshold concentration and over a narrow range of
concentrations53. However, the evolution of an ability to detect
a certain threshold concentration would depend on the absolute amounts available
in nature, and thus a specific threshold (low or high) is not a valid indicator of the
distance over which a semiochemical functions. Still, the breadth of the dose-
response range (often shown to cover 5 orders of magnitude) does appear to be
a valid indicator of a compound's role in distance orientation since a wider response
to concentrations would be required over a wider range of distances from the
source53. Since (S)-(-)-cis-verbenol had an effect on
receptor cell response over a wide range, it was suggested to function in long-range
attraction in I. typographus, while MB had a narrow range (two orders of
magnitude) and was thought to be a close-range signal53. Field
tests with various ratios of these two components were reported to support the
concept121. However, when either component was
released alone there was no significant difference in catch on traps 3 m from the
source and few beetles were attracted121. Holding
either component constant and varying the other caused a synergistic increase in
attraction at the source121. Thus the theory of dose-
response range width as an indicator of orientation range function needs more
investigation - especially since the responses to MB were not tested at higher
concentrations which might saturate the acceptors53.
A possible complication when attempting to ascribe a short- or long-range
function to a semiochemical is that receptor response to a wide range of dosages
may also have evolved due to the natural variation in release rates of the
semiochemical. MB synthesis is under endogenous control in male I.
typographus and relatively large amounts are found in the gut (up to 4 ug/gut)
relative to cis-
verbenol16,17. Thus it may
be expected to vary less than cis-verbenol which is dependant on the
quantity of (-)-alpha-pinene75, a function of genetics and vigor
of the tree17 in addition to the beetle's enzymes. The ability to
respond to a wide range of concentrations would be required not only for
orientation, where concentration decreases with distance93, but
also because aggregations of beetles vary in size and pheromone release
rates37.
A third possible factor complicating the interpretation of dose-response curves is
that not all semiochemicals are equally volatile. MB is one of the most volatile of
bark beetle pheromones and is found in one of the largest amounts per beetle
(x=0.5 ug/male), while cis-verbenol is produced in small amounts (0.06
ug/male) and is less volatile17. E,Z-MD is one of the least
volatile of bark beetle pheromone components (having a volatility comparable to
moth pheromones) and is produced in the smallest of amounts (0.01
ug/male)46. Whether this quantitative-volatility relationship holds
more generally for bark beetle pheromones is not yet clear.
