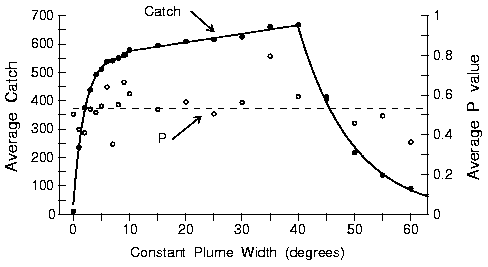
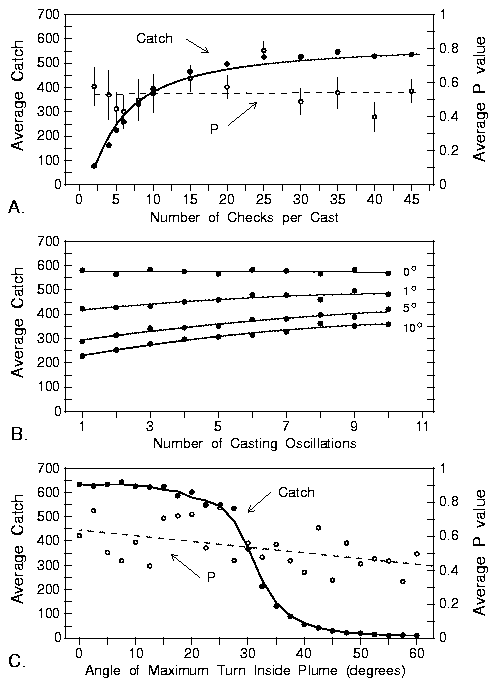
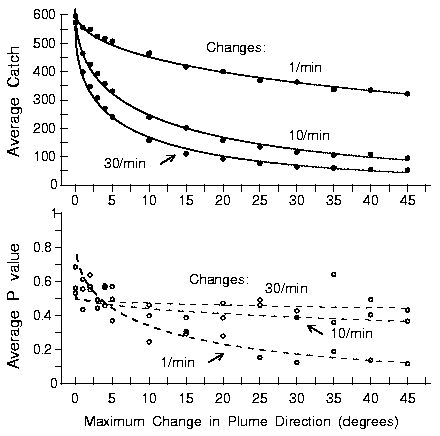
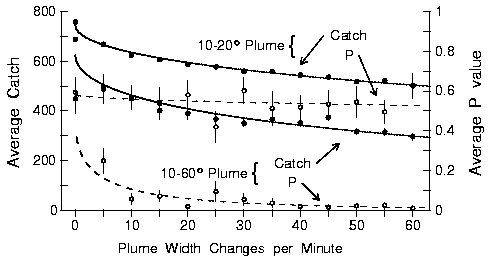
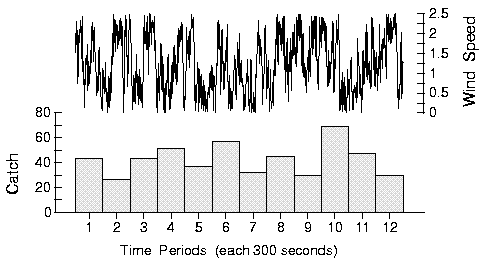
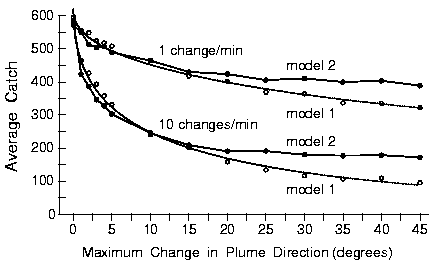
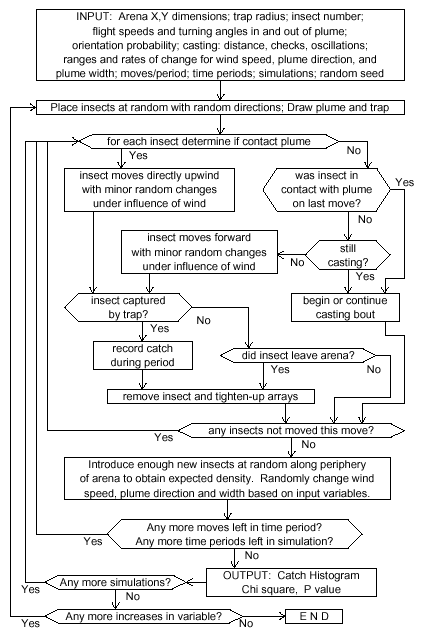
|
|
Present address: 
|
|---|
|
|
Present address:
|
|---|

 Portable fraction collector and pipe trap for catching bark beetles.
Portable fraction collector and pipe trap for catching bark beetles.
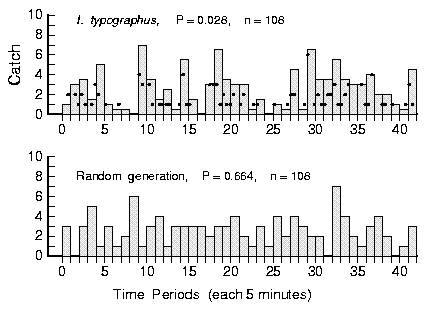
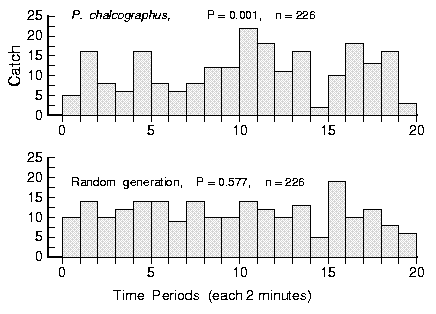
| Table 1. Chi-square analysis of bark beetle arrivals at pheromone-baited traps in the field. The arrival patterns were partitioned into various histograms of different cell widths (1 to 30 min) and compared to average expected frequencies based on total catches using chi-square analysis. P values less than 0.05 indicate that a distribution of catch is not uniformly random. | ||||||||
| Minutes per Histogram Cell | ||||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 4 | 5 | 6 | 10 | 20 | 30 | |
| Ips typographus | ||||||||
| May 29, 1984 (14:30) | ||||||||
| df | - | 104 | 51 | 41 | 34 | 20 | 9 | 6 |
| P value | - | 0.005 | 0.004 | 0.028 | 0.054 | 0.326 | 0.518 | 0.607 |
| June 5, 1984 (14:30) | ||||||||
| df | - | 29 | 14 | 11 | 9 | 5 | 2 | 1 |
| P value | - | 0.004 | 0.002 | 0.063 | 0.007 | 0.310 | 0.290 | 0.450 |
| Pityogenes chalcographus | ||||||||
| June 16, 1987 (14:30) | ||||||||
| df | - | 19 | 9 | 7 | 5 | 3 | 1 | - |
| P value | - | < 0.001 | < 0.001 | 0.016 | 0.013 | 0.141 | 0.033 | - |
| June 16, 1987 (15:18) | ||||||||
| df | 19 | 9 | 4 | 3 | 2 | 1 | - | - |
| P value | < 0.001 | < 0.001 | 0.002 | < 0.001 | 0.002 | 0.003 | - | - |
| June 17, 1987 (14:46) | ||||||||
| df | 19 | 9 | 4 | 3 | 2 | 1 | - | - |
| P value | 0.009 | 0.134 | 0.015 | 0.018 | 0.068 | 0.222 | - | - |